









au sommaire
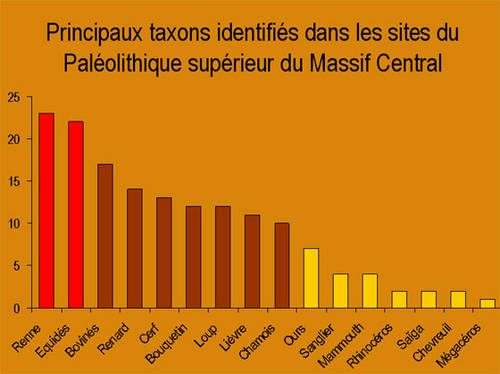
Au Paléolithique supérieur, les taxonstaxons représentés sont les mêmes que ceux des autres régions de France et on peut les classer en trois groupes. Les gibiers présents dans quasiment tous les sites sont le rennerenne et le cheval.
 dans son habitat naturel. © Richard Bartz, <em>Wikimedia commons</em>, CC by-sa 2.5")

Les plus rares correspondent soit à des herbivoresherbivores mieux représentés dans les périodes anciennes (mammouth, rhinocérosrhinocéros et mégacéros), soit à des herbivores qui se développeront à l'HolocèneHolocène et qui traduisent des climatsclimats plus tempérés (sangliersanglier et chevreuilchevreuil), à deux exceptions près. En effet, le saïgasaïga reste rare, peut-être en raison de la faible représentation des périodes qui ont connu son développement, et l'ours est un cas un peu particulier, son statut de gibier étant rarement démontré dans ces sites.


Des espèces communément chassées
Le troisième groupe comprend les gibiers représentés dans environ la moitié des sites. Il s'agit des autres herbivores communément chassés (bovinés, cerf, bouquetinbouquetin, chamoischamois, lièvre) et des carnivorescarnivores les mieux représentés (loup et renard) (3).


Enfin, la répartition des espècesespèces chassées ne présente aucune particularité Nord/Sud à l'exception de la présence, pour l'instant exclusive (mais si faible), des gros herbivores (mammouthmammouth, rhinocéros, mégacéros) dans le Nord puisqu'elle est liée aux sites anciens sous abris de Teilhat et de Châtelperron (cf. figure 1).

Les données concernant l'Épipaléolithique et le Mésolithique proviennent de sept sites et il n'est pas encore certain que la première phase de la recomposition faunique, l'Alleröd, soit documentée. En effet, un certain nombre de corpus issus de fouilles anciennes et attribués à un Magdalénien dit « terminal » posent problème. C'est au moins le cas de Béraud, de Sainte-Anne 2, de Tatevin et de BaumeBaume-Loire. Il nous semble clair que les corpus fauniques de Béraud et de Sainte-Anne 2 sont postérieurs à 12 300/12 200 BP (absence du renne et présence d'espèces forestières marqueurs : Fontana, inédit), ce qui reste à confirmer par la datation radiocarbone des espèces marqueurs et de l'industrie osseuse associée, issues des niveaux et secteurs appropriés.

À Tatevin, le renne est représenté et doit donc être daté de toute urgence afin de vérifier l'antériorité de cette occupation par rapport aux précédentes. Enfin, le cas de Baume-Loire est encore plus problématique puisque J. Bouchud a identifié, en 1966, le renne dans un cortège faunique qu'il attribue (sur les bases de la microfaune et de l'avifauneavifaune) au DryasDryas III, attribution identique à celle que fera A. Crémillieux près de 10 ans plus tard sur la base de l'industrie lithique épipaléolithique et d'une date radiocarbone. Or, la présence du renne au Dryas III n'est aujourd'hui pas documentée dans la seule région qui possède des données issues de datations radiocarbone.

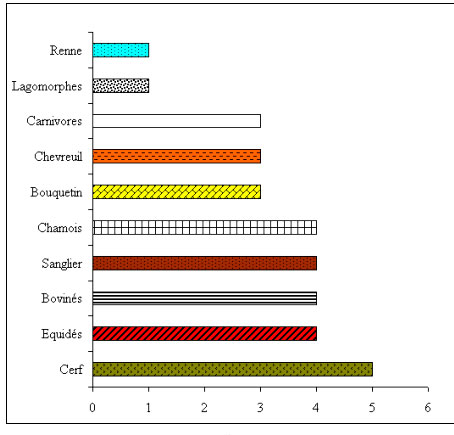
Notre documentation pour la période 12 000 - 10 000 BP est donc très limitée et nous permet seulement de constater qu'en 10 000 BP, les espèces qui vont se développer durant l'Holocène sont déjà présentes et que leur nombre est, comme partout en France, plus réduit (figure 4).
Quant aux restes de micromammifères et d'oiseaux, ils ont rarement été étudiés. Seules les publications de J. Chaline, de J.-C. Marquet, de J. et P. Bouchud et de C. Mourer-Chauviré (4) ont fourni quelques données concernant moins de dix sites. De la même façon, les données concernant l'ichtyofaune se limitent à quelques mentions éparses, une étude sous presse et une étude en cours.
3 - L'origine, anthropique ou naturelle, des restes de ces deux canidéscanidés est rarement discutée.
4 - Ainsi qu'une étude en cours de M. Jeannet et C. Mourer-Chauviré (Fontana et al., 2003)












. © John H. Ghent, Wikimedia commons, CC by-nc 3.0")
. © Diliff, CC By 2.5")


, responsables de quelque 40 000 décès en France. © Ponchita, Adobe Stock")




")


