









au sommaire
La rétinerétine possède une structure complexe dans laquelle on peut reconnaître une dizaine de couches. Quels sont les rôles des cônescônes et des bâtonnetsbâtonnets ? Comment ces photorécepteurs de nos yeuxyeux envoient-ils des messages au cerveaucerveau ?



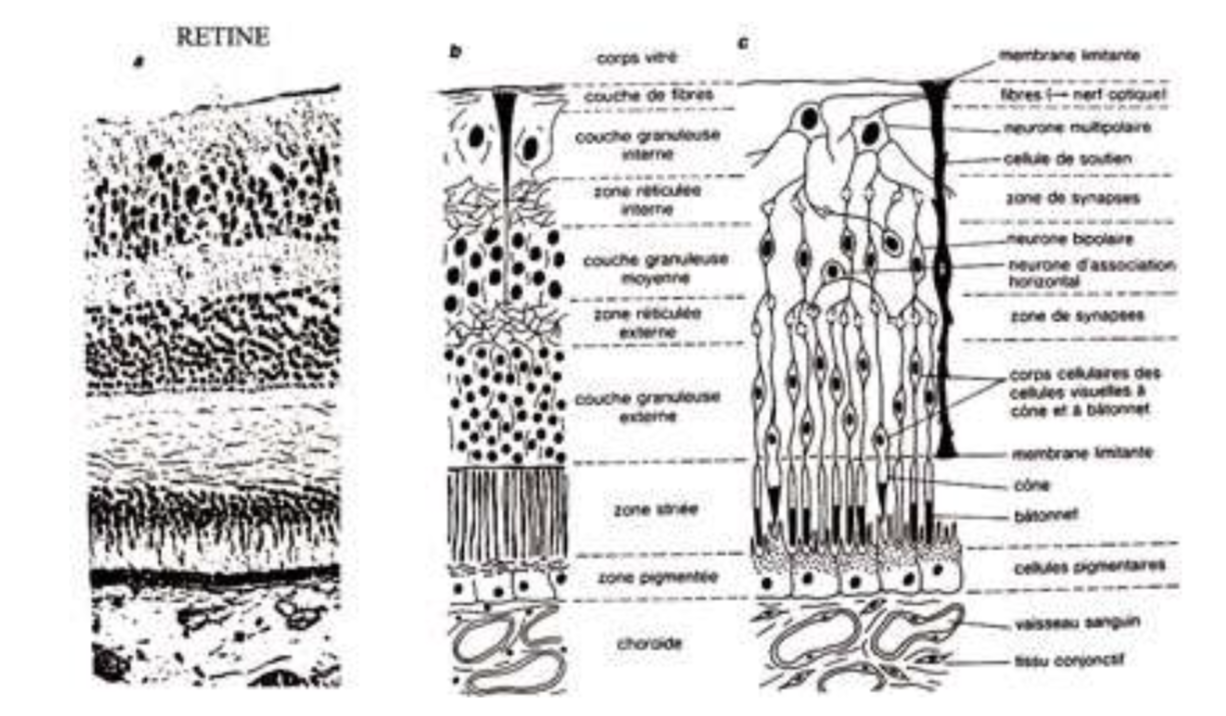
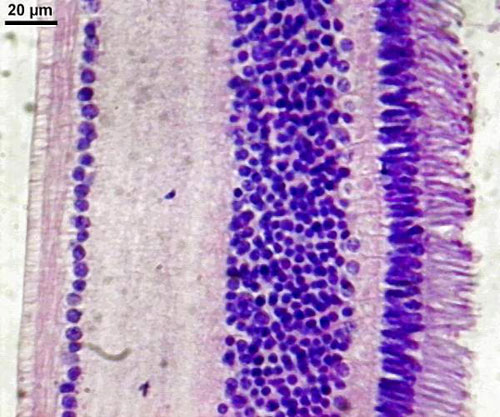
Les différentes couches de la rétine
- La couche la plus externe est adhérente à la choroïdechoroïde.
- La couche des cônes et des bâtonnets est formée par les deux segments externes et internes des cellules visuelles : c'est là que commencent les premiers phénomènes de la vision. Elle est épaisse de 4 µm et compte environ 130 millions de bâtonnets et 65 millions de cônes.
- La limitante externe est une membrane.
- La couche des grains externes est constituée par les corps cellulaires des cônes et des bâtonnets.
- La plexiforme externe, épaisse de 20 µm, est formée par la jonction synaptique des cônes et des bâtonnets avec les dentrites des cellules de la couche suivante. Elle est le siège du relais entre rétine sensorielle et rétine cérébrale.
- La couche des grains internes, épaisse d'environs 30 µm, est constituée par les neurones bipolairesneurones bipolaires qui transmettent l'influx nerveux de la cellule réceptrice à la cellule ganglionnaire, par les cellules d'association et les cellules de soutien : corps cellulaire des fibres de Müller, cellules amacrines.
- La plexiforme interne, où se fait la jonction cellules bipolaires - cellules ganglionnaires, est épaisse de 20 à 30 µm.
- La couche des cellules ganglionnaires est faite de grosses cellules nerveuses, dont les cylindraxes, très longs, constitueront le nerfnerf optique. Épaisse de 10 à 20 µm, elle est presque partout faite d'une seule couche, sauf autour de la fovéafovéa, où les noyaux s'empilent sur 7 à 8 rangs expliquant le relief observé à ce niveau.
- La couche des fibres optiquesfibres optiques se dirigeant vers la papille optique.
- Enfin, la limitante externe est formée par la réunion des extrémités internes des fibres des cellules de Müller (rôle de soutien).
Chacun des types cellulaires, cônes et bâtonnets, de ces couches possède une répartition et des caractéristiques physiologiques qui leur sont propres.

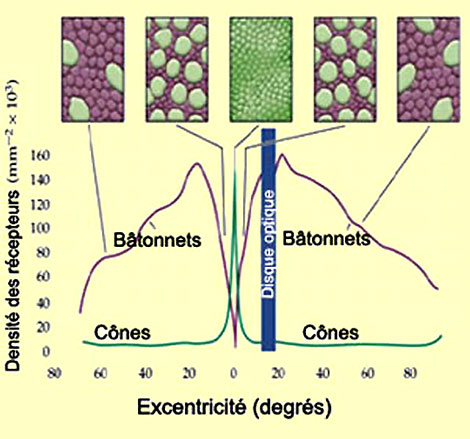
La répartition des cônes et des bâtonnets n'est pas uniforme sur la rétine, les bâtonnets étant beaucoup plus nombreux dans la rétine périphérique et les cônes dans la fovéa. Le nombre de photorécepteurs connectés à une même cellule ganglionnaire est aussi beaucoup plus grand en périphérie. L'effet combiné de cette organisation est d'accroître la sensibilité à la lumière en périphérie de la rétine. La contrepartie est que la précision de l'image souffre de la convergence de nombreux photorécepteurs sur une même cellule ganglionnaire.

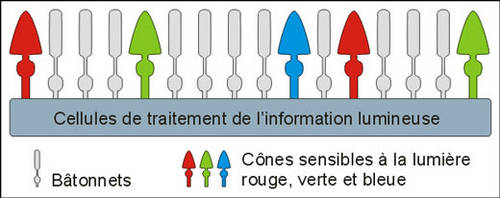
Les cônes fonctionnent avec la lumière
Le segment interne contient le noyau et les organitesorganites (les mitochondriesmitochondries, l'appareil de Golgiappareil de Golgi, etc.)) indispensables au fonctionnement de toute cellule. Dans sa partie la plus distale, son pied, de taille relativement grande, le segment interne possède divers types de synapsessynapses : des synapses électriques, où le transfert du message nerveux est comparable à une simple conduction électrique, assurent des relations entre photorécepteurs voisins (cônes et bâtonnets), et des synapses chimiques qui permettent la transmission du message nerveux à l'aide d'un neuromédiateur ; il s'agit dans le cas présent d'un acide aminéacide aminé : le glutamateglutamate. Celui-ci est contenu dans le bouton synaptique (renflement du prolongement pré-synaptique) et sera expulsé de la membrane pré-synaptique à la membrane post-synaptique déclenchant ainsi un influx nerveux.

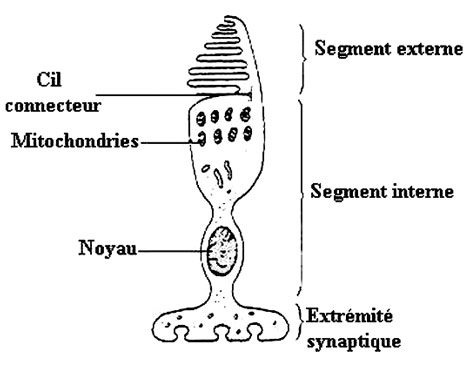
L'autre partie du photorécepteur est son segment externe. Il est formé de l'empilement de plusieurs centaines de lamelles, elles-mêmes correspondant à des repliements de la membrane plasmiquemembrane plasmique qui enveloppe le photorécepteur. C'est au niveau de cette partie du cône, le segment externe, que se produit l'interaction avec la lumière. Les molécules impliquées, les iodopsines, sont présentes dans la membrane des lamelles.
Le système des cônes est un système à haute résolutionrésolution mais sa sensibilité est limitée. En 1967, un biologiste nommé Tornita enregistre l'activité électrique de 142 cônes de carpecarpe ; ceux-ci sont éclairés par des éclairséclairs de lumière monochromatique dont il fait varier la longueur d'onde entre 400 nm (violet) et 700 nm (rouge). Il remarque que certains cônes ont une réaction électrique maximale différente. Ces résultats semblent indiquer l'existence de trois sortes de cônes ayant une absorptionabsorption maximale, une dans le bleu-violet, à 420 nanomètres, la deuxième dans le vert, à 530 nm, et la troisième à 565 nm, dans le jaune-rouge. Ces résultats seront confirmés par l'extraction de trois sortes de pigments des cônes de rétines humaines. Ces spécificités ont permis de classer les cônes humains en trois catégories :
- les cônes « S », Short, qui contiennent en majorité le pigment sensible au bleu ;
- les cônes «M », Medium, présentant une concentration plus importante en pigments sensibles au vert ;
- et enfin les cônes « L », Long, porteurs du pigment sensible au rouge.

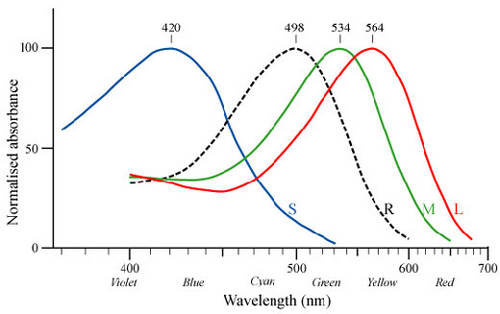
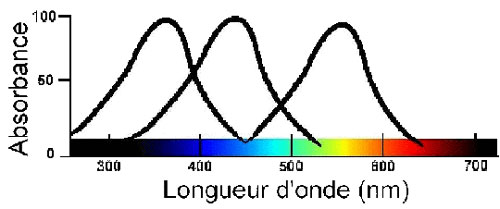
À titre de comparaison, voici le même graphique concernant la sensibilité à la lumière chez les insectesinsectes.

Les bâtonnets ne détectent pas la couleur
Ce sont des cellules du même type - mais de forme différente - des cônes. Elles ne sont sensibles qu'à la quantité de lumière et sont utilisées pour la vision nocturnenocturne.

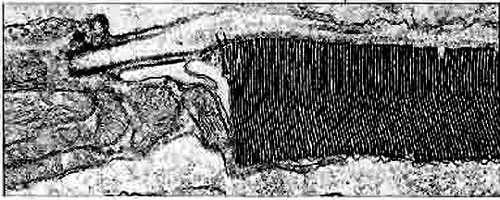
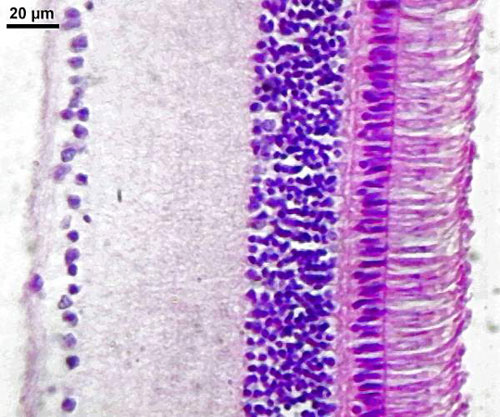
Pour bien se rendre compte de la différence de vision entre la vision diurnediurne des couleurscouleurs avec les cônes principalement et la vision nocturne des valeurs avec les bâtonnets, on peut comparer les rétines des oiseaux diurnes et nocturnes : on y voit bien la différence de nombre de photorécepteurs, essentiellement des bâtonnets pour les nocturnes.


La photoexcitation et la lumière
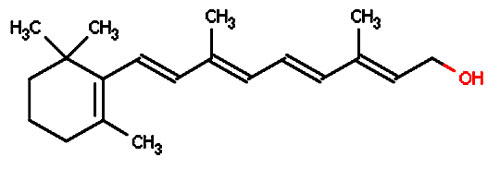
Une protéineprotéine transmembranaire, l'opsine, sur laquelle vient se fixer un groupement le rétinène, qui n'est autre qu'un aldéhydealdéhyde de la vitamine Avitamine A ou rétinol subit l'arrivée d'un photonphoton entre 650 et 400 nm sur la partie rétinène de la rhodopsinerhodopsine qui passe alors de la conformationconformation 11-cis à la conformation tout-trans (appelée métarhodopsine II).

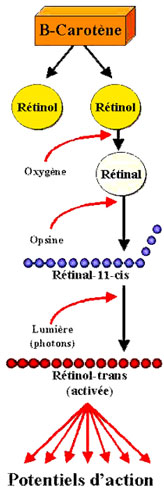
Le rétinol est issu de la pro-vitamine A ou bêtacarotène, fournie par l'alimentation : carottescarottes, abricotsabricots, myrtillesmyrtilles, beurre, épinardsépinards. Cette transformation a deux effets :
- le rétinène trans se détache spontanément de la molécule d'opsine, il diffuse dans le milieu intra, puis extracellulaire où il est repris et, sous action enzymatiqueenzymatique il est reconverti en rétinène 11-cis, au bout de 40 à 60 minutes chez l'homme ;

- la métarhodopsine II produite par transformation de la rhodopsine sous l'effet de la lumière active une protéine : la transducine. Par la suite, les canaux Na+ des cellules réceptrices se fermentferment, provoquant une hyper-polarisation membranaire qui engendre un potentiel d'actionpotentiel d'action électrique dans les cellules ganglionnaires.

Reproduction et utilisation interdites
Le fonctionnement des cellules photosensibles consiste donc à transformer de l'énergieénergie lumineuse en énergie électrique, qui elle sera gérée par les neurones des centres cérébraux pour former une image compréhensible pour l'observateur.
La transmission de l'information
Le message nerveux est acheminé jusqu'au cerveau, il est transmis d'une cellule nerveuse à une autre. Le fonctionnement d'une synapse qui sert de zone de contact entre les deux neurones est complexe. L'extrémité du prolongement pré-synaptique est formé d'une grosseur, le bouton synaptique, riche en neurotransmetteursneurotransmetteurs contenus dans de petites vésicules. Il existe un espace séparant la zone pré-synaptique de la zone post-synaptique : la fente synaptiquefente synaptique. La membrane post-synaptique, censée recevoir l'influx, porteporte des récepteurs spécifiques à un neurotransmetteur et lorsqu'un un influx nerveux atteint le bouton synaptique, il y provoque l'expulsion du neuromédiateur dans la fente par éclatement des vésicules. Celui-ci atteint alors les sites récepteurs de la membrane post-synaptique et y déclenche un influx nerveux. Dans le cas étudié, le glutamate, libéré par les cônes, nécessite un certain seuil de dépolarisation pour que les vésicules éclatent. Cette dépolarisation correspond à une modification du potentiel de récepteur mesurable de part et d'autre de la membrane. La quantité libérée augmentera alors en fonction du niveau de dépolarisation.
. 4. Libération du neurotransmetteur, ici le glutamate. 5. Fixation par le récepteur. 6. Nouveau potentiel dans le neurone suivant. 7. désactivation enzymatique du transmetteur. 8. Recapture (endocytose) et le cycle recommence... © Reproduction et utilisation interdites")
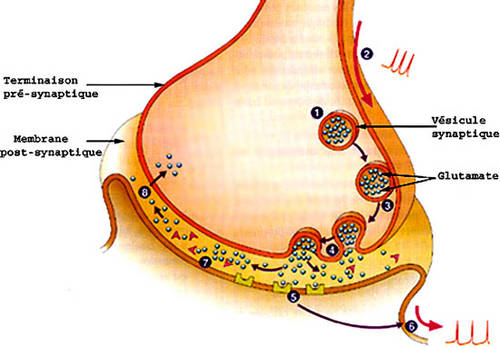
À côté de cette voie directe qui va des photorécepteurs au cerveau, deux autres types de cellules participent au traitement de l'information visuelle dans la rétine. D'une part, les cellules horizontales reçoivent de l'information des photorécepteurs et la transmettent à plusieurs neurones bipolaires environnants. Et d'autre part, les cellules amacrines reçoivent leurs inputs des cellules bipolaires et activent les neurones ganglionnaires dans les environs. Les neurones de la rétine « couvrent » chacun une région de notre champ visuelchamp visuel. Cette région de l'espace où la présence d'un stimulus approprié modifie l'activité nerveuse d'un neurone est appelée le champ récepteur de ce neurone.
Le traitement des images par le cerveau
Les voies de la sensibilité visuelle consciente, partiellement croisées dans le chiasma, aboutissent dans la région occipitaleoccipitale. Toute lésion de l'aire visuelle entraîne une cécité partielle correspondant à une région définie du champ visuel. La rétine se projette point par point sur l'aire visuelle, mais le territoire cortical correspondant à la fovéa est relativement très étendu.
Les neurones du cortexcortex visuel primaire ont, quant à eux, des champs récepteurs plutôt allongés. Ils répondent bien à des traits de lumière ayant une orientation spécifique. Ces champs récepteurs répondant à une orientation donnée sont ceux des cellules simples. Ces champs récepteurs de forme rectangulaire ont souvent une bande centrale répondant positivement à la lumière (ON) flanquée de deux bandes répondant à l'obscurité (OFF). Si le trait de lumière n'est pas sur la bande ON, le stimulus n'est tout simplement pas transmis.

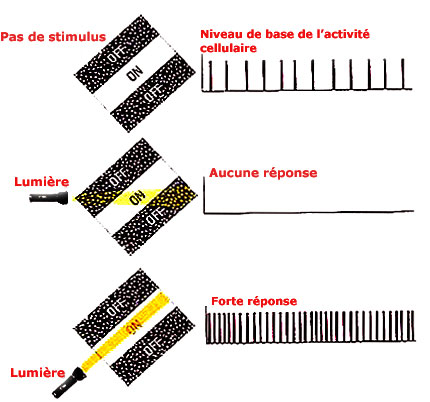
D'autres cellules du cortex visuel primaire ont des champs récepteurs dits complexes et hypercomplexes qui ont des propriétés encore plus sélectives.
L'aire visuelle du cerveau
Elle est divisée en plusieurs régions dont on ne connaît pas tout.
- L'aire V1 et V2 : ces aires jouent un rôle très important dans la perception des contours. Elles restent incontournables dans toute perception visuelle fine.
- L'aire V3 : les scientifiques n'ont pas clairement identifié cette aire.
- L'aire V4 : cette aire joue un rôle dans la perception des couleurs comme l'a démontré Semir Zeki de l'University College de Londres. Mais les spécialistes pensent que d'autres régions y sont associées.
- L'aire V5 : elle joue un rôle dans la perception des mouvementsmouvements comme le montrent des expériences par imagerie à résonance magnétique.
L'aire psycho-visuelle
En avant de l'aire visuelle primaire s'étend l'aire psycho-visuelle. La stimulationstimulation électrique d'un point de cette aire fait apparaître des hallucinationshallucinations évoquant des objets ou même des scènes plus complexes. La destruction partielle de l'aire secondaire entraîne une agnosie visuelle : le sujet voit les objets, mais ne les reconnaît pas ; il a perdu tout souvenir des perceptions antérieures. C'est le cas par exemple dans la cécité verbale : le malade voit parfaitement la page écrite placée sous ses yeux, mais il ne peut en déchiffrer un mot, comme si cette page était composée de signes inconnus.
La destruction des lobes occipitaux crée une « nuit psychique » beaucoup plus complète qu'en cas de lésion sur les voies optiques : non seulement le sujet est aveugle, mais il a perdu tout souvenir de ses perceptions visuelles antérieures.
Et puis l'information peut être retraitée dans les lobes frontaux. Ce chapitre concerne la neurophysiologie cérébrale, nous sortons du cadre de cet exposé.


")



 ou de rétinopathies pigmentaire. © MS Photographie, Fotolia")



. Un femtomètre vaut 10-15 = 0,000 000 000 000 001 mètre. © Jean-Paul Ebran, CEA")
