




au sommaire
Nous allons maintenant étudier certains macrofossiles d'invertébrésinvertébrés et caractériser leur évolution. En effet, selon les espècesespèces, la morphologiemorphologie évolutive change et des critères apparaissent ou disparaissent.


Fossile Cardiidae © PalaeoMal - CC BY-SA 3.0 et GFDL
1. Les Lamellibranches (ou bivalves)


Les Lamellibranches représentent une dégénérescence du plan mollusquemollusque : il s'agit d'une simplification évolutive (réversionréversion). En effet, ils se sont adaptés à un mode de vie sédentaire. Ils peuvent évoluer en conservant une forme larvaire à l'âge adulte (néotenie). Dans ce cas, le pied change de fonction et les branchies peuvent être remplacés par des poumonspoumons : c'est une adaptation à la vie terrestre (ex : Salamandres). Au niveau des muscles, les Lamellibranches ont évolués d'une forme isomyaire (deux muscles de taille identique) vers une forme anisomyaire (deux muscles de tailles différentes) puis monomyaire (un seul muscle) : c'est dans ce cas une évolution par simplification. Les dents disparaissent chez les formes à habitat très protecteur (Lamellibranches perforants ou fouisseurs) comme les huîtres. La symétrie est aussi affectée par l'évolution. En effet, d'une double symétrie (parallèle et perpendiculaire aux valves), on arrive à une symétrie unique parallèle aux valves issue d'une réduction d'un des bords puis vers une dissymétrie totale avec perte d'un muscle. Cette évolution de symétries est couplée avec la perte des dents. La radiation adaptativeradiation adaptative des Lamellibranches est donc la suivante. A partir d'une forme taxodonte (dents nombreuses indifférenciées), on passe à une forme hétérodonte (dents inégales peu nombreuses différentiées) par une évolution simplifiante liée au nombre de dents et complexifiante au niveau de la forme des dents. Puis, de cette forme hétérodonte, on arrive à une forme schizodonte (dents fendues en deux) et pachyodonte (dents énormes et épaisses). Parallèlement à cette lignée, on observe à partir des taxodontes une évolution vers de pseudotaxodontes (beaucoup de dents asymétriquesasymétriques) et vers les isodontes (dents de même taille de part et d'autre du ligament) ou les huîtres (plus de dent). Mais ce qu'il faut retenir de l'évolution des Lamellibranches est cette simplification évolutive par perte de caractères : la réversion.
2. Les Gastéropodes
Du point de vue évolutif, les Gastéropodes ont été soumis à une évolution qui a eu lieu au stade larvaire. A ce stade, la cavité maléale est amenée en position frontale par une flexure de 180°. Le mollusque est donc devenu asymétrique. Les formes les plus anciennes, les archéoGastéropodes sont proches des Monoplacophores (ancêtre communancêtre commun à tous les mollusques). Ils évoluent vers les prosobranches (branchies vers l'avant) puis vers les monobranches (une seule branchie). Ils conquissent les continents par vaguesvagues successives (cladesclades) en transformant leur branchie restante en poumon. Ces vagues ont lieu au CarbonifèreCarbonifère, JurassiqueJurassique, CrétacéCrétacé et à l'EocèneEocène. Ils s'adaptent d'abord aux eaux douces puis à la vie terrestre. Au Crétacé, la vague est constituée des Helicidae (escargots). Les opistobranches évoluent vers une forme terrestre au Tertiaire. Parallèlement à cette évolution par conquête des continents en perdant une branchie puis en transformant la branchie restante en poumon, des Gastéropodes initialement herbivoresherbivores deviennent carnivorescarnivores, c'est le cas des Coengast.
3. Les Céphalopodes

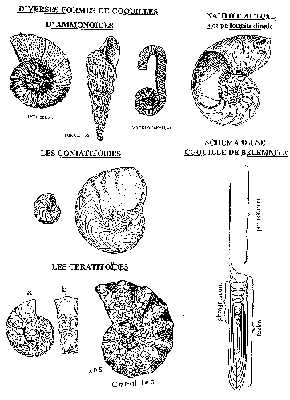
L'ancêtre des Céphalopodes est le Monoplacophore. L'évolution s'est faite dans le sens où la coquille s'est refermée et les cloisons sont apparues. A l'origine, il existe trois types de coquille : droites (orthocône), arquées (cystocône) ou enroulées (nautilocône, origine des Nautiloïdes). Ces formes ont évolué au cours du temps puisque la meilleure façon de nager est d'être enroulé. En effet, le centre de gravité de l'animal est au centre de la coquille. Parallèlement à la lignée des Nautiloïdes, les Bactritoïdes sont à l'origine des Colleoïdes et des Ammonoïdes, on dit que c'est un groupe charnière. L'innovation principale des Bactritoïdes est la création de formes enroulées (évolutes et involutes). Il existe toutes les formes intermédiaires au DévonienDévonien. A partir des Bactritoïdes, les Ammonoïdes représentent le plus grand groupe en variété et le succès du type enroulé. L'évolution des Nautiloïdes se fait par complexification du système de cloisons et adaptation aux milieux plus profonds (la coquille est plus solide en s'allongeant). En effet, du stade Goniatite (cloison en zigzag), on passe au stade Cératite (selles et lobes qui s'allongent et se replient) puis au stade AmmoniteAmmonite (sutures persillée plus complexe). Ce passage se fait par un système de ramification : un seul genre passe une crise et on observe alors une alternance de disparition apparition. C'est la radiation adaptative qui correspond à la diversification à partir d'un groupe (exploitation des niches écologiques libérées lors d'une crise). Cependant, certaines formes déroulées apparaissent : c'est dans ce cas une réversion (groupe des Lytoceratides). Les Colliloïdes, issus des Bactritoïdes ont passé la crise crétacé tertiaire. Leur évolution est basée sur la réduction du rostre, l'internalisation de la coquille, la réduction du phragmocône puis la disparition de la coquille. L'innovation majeure des Céphalopodes est donc la création de l'appareil hydrostatique et son perfectionnement couplée avec les formes enroulées plus propices à la nage et aux grandes profondeurs avec le développement de cloisons persillées. De plus, ils ont concentré leur système nerveux à l'avant de l'animal.
4. Les Brachiopodes

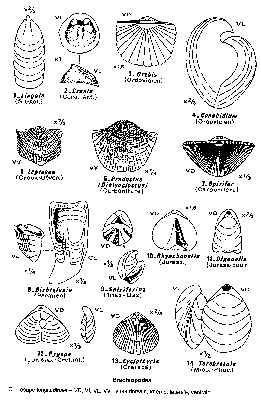
Les principaux changements relevés chez les Brachiopodes nous montrent que la morphologie de la coquille évolue et que l'appareil brachial tend à se complexifier chez la plupart des Brachiopodes. Les Brachiopodes ont évolués de manière quantique. L'effectif de l'espèce était initialement grand. Une crise a fortement réduit cet effectif et la libération de niches écologiques a engendré l'essor d'une nouvelle espèce. En effet, par consanguinitéconsanguinité, l'espèce a pu évoluer très rapidement et l'espèce finale est différente de l'espèce initiale. Il existe désormais entre ces espèces une barrière d'interfécondité. Dans certaines familles de Brachiopodes, on note des clades. Nous avons donc au sein de ces Brachiopodes deux sortes d'évolution : l'évolution anagénétique quantique et l'évolution cladogénétique quantique.
5. Les Arthropodes (Trilobites)

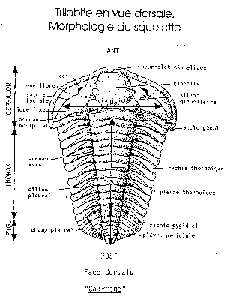
On note une première radiation adaptative des Trilobites au Cambrien inférieur et une seconde au Cambrien moyen qui correspond à l'adaptation à la vie sur la plate-forme (transgression marinetransgression marine). Puis, à l'OrdovicienOrdovicien, le déclin des Trilobites commence, malgré l'émergenceémergence de formes enroulées capables de se protéger, avant une extinctionextinction définitive au PermienPermien. L'évolution des ArthropodesArthropodes s'est faite par perte de cellules visuelles et vers des formes aveugles vivant dans les grands fonds des océans (moins de prédateur que sur la plate-forme). Des formes avec des épines en plus des pattes sont par ailleurs mieux adaptées à la vie sur les récifs. En plus de ce caractère, les Trilobites vont acquérir une suture propariale et la glabelle va se dilater. La disparition de ces récifs lors de la crise Permier/TriasTrias éliminera définitivement les Trilobites. Mais il existe d'autres Arthropodes comme des insectesinsectes ou crustacéscrustacés qui sont encore des formes actuelles.
6. Les Echinodermes

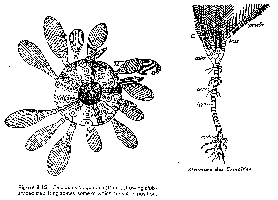
Les formes d'Echinodermes ont évolué en fonction du lieu de vie : les pics que certains possèdent seront larges sur des récifs alors qu'ils seront fins sur milieu vaseux. Le nombre de pores diminue en fonction du nombre de plaque. Les formes libres évoluent vers des formes en cloches puis plates dans des milieux peu profonds. La bouche a migrée vers l'avant et disparaît ensuite.
Mais le moteur de l'évolution est l'alternance de crises et périodes de prospérité. En effet, une crise provoque la disparition de nombreuses espèces et libère des niches écologiques qui serviront à l'essor des espèces qui ont réussies à passer cette crise. De plus, les nombreuses crises ont été propices aux carnivores puisqu'ils ne sont pas dépendant du phytoplanctonphytoplancton largement touché par ces crises répétées. Ceci explique le paradoxe de la prolifération des formes horizontales.

par Sébastien Potot & Julien Bertholon
le 25 octobre 2001


 ont hérité de 1 à 4 % de leurs gènes de l’Homme de Neandertal. Des métissages se sont donc probablement produits dans la population ancestrale des non-Africains issus des régions du Levant et de l’Afrique du Nord. Aujourd'hui, les scientifiques ont également découvert que les Dénisoviens ont légué de 4 à 6 % de leur matériel génétique aux Mélanésiens. Sur ce schéma, les flèches (notées f) indiquent les transferts successifs de gènes entre Néandertaliens (Neandertal), Denisoviens (Denisova) et Mélanésiens (Melanesian). N représente la taille effective des populations, t et tGF (sur l'échelle du temps) marquent une séparation entre deux populations et la période où les flux de gènes ont eu lieu. © David Reich, Harvard Medical School")


, Mahdi Zamani (ESA/Hubble)")


")


