







au sommaire
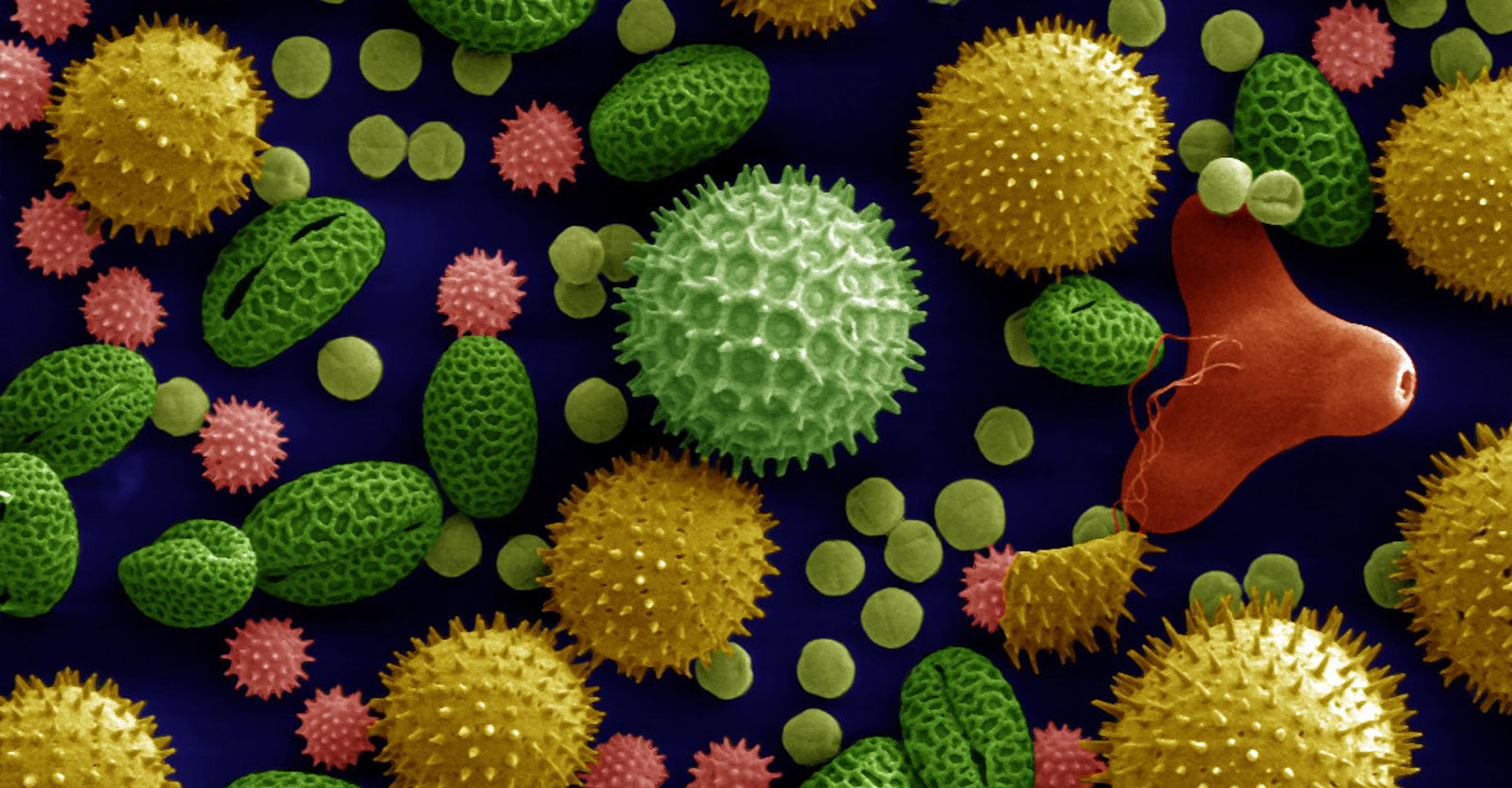
Le pollenpollen de la fleur, partenaire mâle, renferme deux gamètesgamètes ou la cellule qui les génère ; c'est un organisme adapté à la dissémination et à la survie. Il joue un rôle primordial dans la fécondationfécondation chez la fleur.
La teneur en eau du pollen est faible et son métabolismemétabolisme nul. Pour reprendre son activité, il doit se réhydrater à la surface d'un pistilpistil compatible : le stigmatestigmate de la fleur. Il pousse alors un tube ; on dit qu'il germegerme, et assure ainsi le transfert des gamètes au sac embryonnaire, voir photos ci-dessous. Voir également notre dossier « La pollinisation, un service écologique gratuit ».
, lys (<em>Lilium auratum</em>), onagre (Oenothera fruticosa) et Ricin commun (<em>Ricinus communis</em>). ©<em> Dartmouth Electron Microscope Facility, Dartmouth College,</em> DP")
Un contact physique entre le pollen et l'ovule
À la fin du XIXe siècle, Brongniart puis Brown décrivirent l'entrée du tube pollinique dans l'ovuleovule à travers le micropyle et la formation subséquente de l'embryonembryon dans le sac embryonnaire. Ce travail résolut en partie l'énigme du contact pollen-ovule.
En démontrant ce contact physique entre l'ovule et le pollen, ils élevèrent la fécondation chez les plantes au même niveau que celle des animaux, chez qui la nécessité d'un contact physique entre ovule et spermesperme commençait à être acceptée.


Récemment, l'analyse de mutants affectés dans la fécondation a permis de mettre en évidence plusieurs systèmes de signalisation entre tube pollinique et sac embryonnaire, signalisation permettant le guidage du tube vers le micropyle.
Strasburger décrivit la première fécondation chez les fleurs en 1878 ; quelques années plus tard, S. Nawashin et L. Guignard décrivirent la double fécondation chez le lys (1898, 1899).


La fécondation in vitro chez les plantes
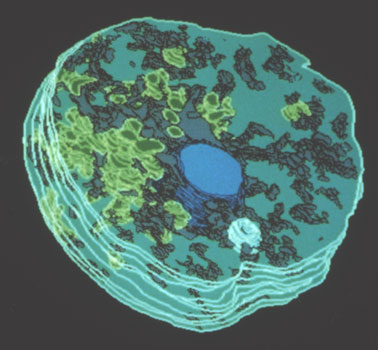
Dans les années 1990, nous avons réussi la première fécondation in vitro chez les plantes et mis en évidence un flux cacique à l'origine de l'activation de l'oosphère et de la formation ultérieure de l'embryon.
Un avantage de ce système était la possibilité de contrôler le site d'adhésion gamète mâle-gamète femelle et de suivre par des techniques appropriées, électrochimiques en particulier, la capacitationcapacitation de l'oosphère (photo ci-dessous). Une telle approche est impossible chez les autres systèmes biologiques animaux ou chez les alguesalgues.

Cette approche est pourtant réductionniste : ce qui est vu in vitro ne correspond peut-être pas à ce qui existe in vivoin vivo. D'où l'intérêt des approches actuelles, notamment à l'aide de mutants qui permettent de sérier les étapes de la fécondation et de caractériser les gènesgènes responsables, donc les mécanismes intimes en jeu.
Ainsi, un signal positif issu de la cellule-œuf entraîne la prolifération de l'albumenalbumen. C'est la caractérisation d'un mutant affecté dans une cyclinecycline CDC2 avec un effet paternel qui serait en cause. Chez ce mutant, le pollen est viable et un seul gamète est fonctionnel, seule la fécondation de l'oosphère a lieu.



















 vient de lancer la compagne « Les records à ne pas battre » pour sensibiliser à l’urgence climatique et à la dégradation de la biodiversité. © Image générée par l’IA Adobe Firefly")
 forment une très vaste famille de coléoptères comprenant plus de 37 000 espèces connues, réparties en 2 500 genres. Même s'il est impossible de le dire en regardant cette image, prise par en dessous, les chrysomèles ont souvent un dos métallisé très coloré. © Chuck Daghlian/Louisa Howard, domaine public")



