






au sommaire


Recherche du consensus dans le génome complet d'A. thaliana
Connaissant maintenant la séquence consensus probable des promoteurs des gènesgènes de la RSARSA, DNADNA-pattern search peut faire la démarche inverse qui consiste à rechercher les promoteurs qui ont plus d'une occurrence de cette boîte et qui sont présents dans le génomegénome complet d'A. thaliana.
En apportant au programme les différentes boîtes qui se rapportent à la RSA, le programme peut rechercher sur le génome complet ces motifs déterminés. Ces séquences sont celles déterminées par les analyses précédentes (tgcagc et gcagc) et celles issus d'études précédentes (ttgac et tgacg). Le résultat de l'analyse montre l'existence de cette boîte consensus dans de nombreux promoteurs (fi-gure 6). Or la grande majorité correspond à des gènes codant des protéinesprotéines inconnues ou des fac-teurs de transcriptiontranscription (tableau 6).
Figure 7. Histogramme représentant le nombre de promoteur en fonction de l'occurrence de chaque boîte
A : consensus déterminé par Oligo-analysis
B : consensus déterminé par Motif Sampler
C : boîte de liaison au facteur de type WRKY
D : boîte de liaison au facteur de type TGA-bZIP
Parmi les protéines dont les fonctions sont supposées, se trouvent des transposases, des protéines de résistancerésistance aux phytopathogènes ou aux stressstress, des perméases, des transporteurs membranaires, des récepteurs membranaires, ...
Dans les protéines enzymatiquesenzymatiques connues, nombreuses sont celles qui agissent dans la réponse aux stress oxydatifsstress oxydatifs ou aux phytopathogènes, ainsi que dans la régulation de la biosynthèse des thio-redoxines. Mais la plupart des promoteurs présentant ce motif régulent des gènes dont la fonction est inconnue comme le sont la majorité des protéines de résistance aux phytopathogènes.
Les gènes de facteurs de transcriptions représentent une part non négligeable dans la répartition du consensus tgcagc dans l'ensemble des promoteurs du génome (tableau 6).

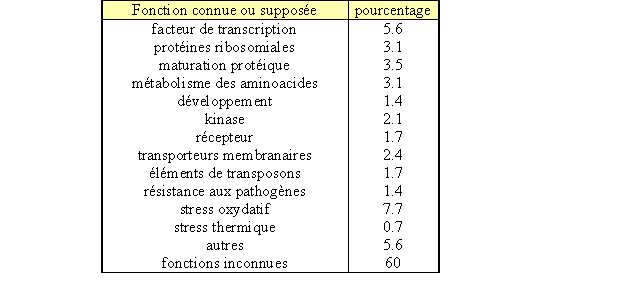
Tableau 6. répartition du consensus tgcagc en fonction du type de promoteur
Les distributions des consensus déterminés par les différentes analyses bioinformatiques sur l'échantillon que sont les promoteurs des gènes de la RSA, peuvent être cumulées comme le montre l'histogramme de la figure 7.

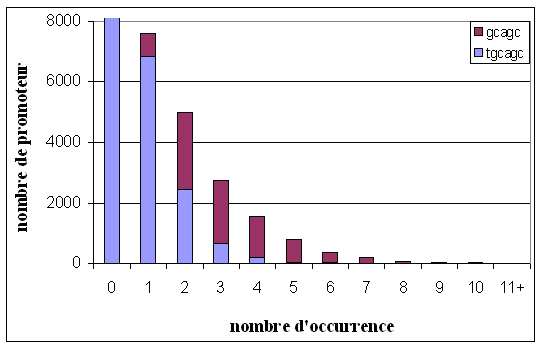
Figure 8. Distribution cumulée des consensus établis par Motif Sampler et Oligo-analysis
La visualisation de la distribution cumulée permet de voir la baisse exponentielle du nombre de promoteur lorsque le nombre d'occurrence augmente. La forme pentamèrique GCAGC se trouve confondue dans celle hexamèrique TGCAGC, tant que cette dernière est présente, car la proportion de promoteur présentant la forme courte par rapport à ceux qui présentent la forme longue, aug-mente au fur et à mesure que le nombre d'occurrence augmente. Or les études de distribution sta-tistique de motif génomiquegénomique, considèrent généralement le seuil de significativité à 3 motifs par promoteur car un ou deux motifs par promoteur peuvent résulter de mutation spontanée sans avoir un sens dans la régulation génomique (Maleck et al., 2000)

par Emmanuel Poulouin
le 25 février 2003






, un ensemble de logiciels pour le séquençage du génome, a été développé au Broad Institute. © Suravid, shutterstock.com")








, au-dessus du lac Svityaz, en Ukraine. © Ihor Khomych, via Spaceweather")






