









au sommaire


Modes de spéciation
1 - Les modes de spéciation
Tout ce que nous avons vu précédemment n'est valide que dans un milieu invariant. Autant dire que c'est une situation uniquement théorique car les milieux changent perpétuellement autant dans leur composition biologique que physique. Dès lors que l'on joue sur les paramètres du milieu il devra nécessairement avoir une adéquation entre l'espèceespèce et son milieu. C'est vraiment la clef de la réussite pour une espèce car la transformation des milieux est généralement rapide et imprévisible. Soit l'espèce a acquis au préalable les capacités d'adaptation nécessaires soit ces transformations lui seront fatales.
Cependant plus l'espèce est capable de coloniser des nouveaux territoires plus son aire de distributionaire de distribution est important et d'une certaine manière plus les individus situés aux extrémités de cette aire deviennent différents entre eux et isolés les uns des autres. En effet, on comprend aisément que la distance puisse devenir un obstacle au transfert de l'information dès lors qu'elle est supérieure aux capacités de transports. Mais l'éloignement s'accompagne généralement en plus de conditions très différentes. Ainsi deux individus d'une même espèce qui sont fort éloignés dans un milieu caractérisé par un gradient peuvent ne plus trouver les conditions nécessaires au transfert de l'information génétiquegénétique. Ainsi au Mali, une espèce largement répandue (ex : Vitellaria paradoxaVitellaria paradoxa) montre un fonctionnement qui rend incompatible le transfert d'information entre les populations situées au Nord et celles situées au Sud. En effet, le gradient climatique se traduit au niveau phénologique par un décalage des périodes de floraison. Les individus situés au Nord sont en fleurs alors que les individus du Sud sont déjà en fruits. On assiste à un isolement génétique des populations. C'est par ce principe d'isolement que la spéciationspéciation intervient, c'est-à-dire que les nouvelles espèces apparaissent. La spéciation résulte de l'isolement reproductif entre deux populations. Globalement on note deux cas distincts de spéciation:
a) la spéciation allopatrique
Adapté d'après Campbell N. A et Reece J. B., 2004 - Biologie - 2ème édition française. De Boeck Université
© Tous droits de reproduction réservés
Une nouvelle espèce naît après qu'une population se soit retrouvée isolée sur le plan géographique de sa population ancestrale. Il y a fragmentation de l'aire de distribution. La transformation des milieux est le principal responsable de la fragmentation des aires de distribution des espèces. Dans notre petit schéma théorique qui montre les allers/retours entre le centre de dispersion d'une espèce et les nouveaux milieux colonisés, nous comprendrons facilement que l'installation d'une barrière autour du centre de dispersion va inhiber le retour des individus migrants et partant le brassage génétiquebrassage génétique des adaptations acquises dans des milieux différents. Les espèces originellement brassées continuent d'évoluer et de s'adapter localement sans ne plus rien s'échanger. On peut comprendre facilement que cette isolationisolation conduit à définir des populations qui ultimement ne seront plus 'compatibles' les unes avec les autres. Dès lors que les deux espèces ne sont plus reproductibles, c'est-à-dire qu'elles ne peuvent plus s'échanger leur patrimoine génétique, il y a apparition d'une nouvelle espèce, cousine de la première mais suffisamment différente pour ne plus être interféconde. Ces barrières géographiques ont plusieurs origines : mouvements des plaques ou modification du cours d'un fleuve, assèchement d'un lac, apparition d'une mer, une faille, une montagne.... glissements de terrain.... Elles correspondent à une transformation d'une partie du milieu qui interdit les échanges entre les deux isolats. L'assèchement d'un lac va empêcher l'échange des espèces hydrochores (qui se disperse par l'eau). La création des océans actuels a défini une barrière géographique (ex: L'atlantique) qui a interdit l'échange génétique entre l'Afrique et l'Amérique du Sud...
b) La spéciation sympatrique

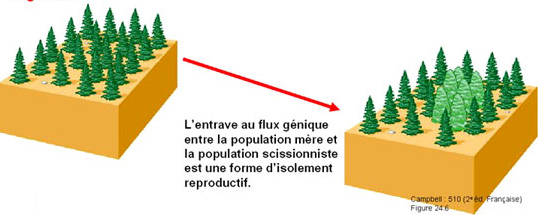
Adapté d'après Campbell N. A et Reece J. B., 2004 - Biologie - 2ème édition française. De Boeck Université
© Tous droits de reproduction réservés
L'isolement reproductif est atteint sans qu'il n'y ait un isolement géographique. Les populations ne sont plus reproductibles entre elles par des processus indirects. Par exemple la disparition ou le changement de régime d'un vecteur responsable de la dispersion ou de la pollinisation aboutit à l'isolement reproductif des deux populations. Dans ce cas, une espèce extrêmement spécialisée dans la dispersion ou la pollinisation par les animaux et surtout si elle a co-évolué avec une espèce animale particulière (ex: Orchidaceae) sera très sensible au changement de comportement de son allié. C'est ainsi que la disparition d'un animal sauvage a des conséquences et des répercutions sur la dispersion de certaines espèces végétales. La création de milieux 'non-naturels' délimite des espaces dans lesquelles les espèces naturelles ne peuvent pas se développer. Les voies de transfert sont théoriquement possibles mais c'est le développement et la croissance des espèces qui devient impossible. Ainsi l'installation d'un village avec les activités humaines associées (champ, parcs, routes, feu,...) va isoler les populations végétales en réduisant (ou éliminant) les contacts entre les individus, en interdisant le passage des vecteurs...Dans cette catégorie on note également la fragmentation du paysage par rapport à un changement climatiquechangement climatique majeur. L'intensification d'un gradient de pluviométrie va se traduire par une polarité dans la distribution des espèces. Les espèces se concentrent autour de leur pôle d'attraction et la majorité des individus situés dans la zone de colonisation va disparaître. Cependant quelques individus vont persister à la faveur de micro-conditions locales relativement plus favorables (au sens relatif). Le retrait des troupes n'est pas intégral, il reste des poches de résistancerésistance ! Ces populations seront isolées dans le territoire par un phénomène de gradient de sélection sans qu'il n'y ait de barrières géographiques au transfert de l'information. De plus, nous l'avons vu, bien souvent, le décalage climatique s'accompagne d'un décalage phénologique qui interdit le croisement entre des populations trop éloignées. Dans ce cas il s'agit d'une barrière physiologique étant donné que les individus d'une même espèce ne sont plus interféconds.
Ces modes de fragmentations s'effectuent à toutes les échelles depuis un massif forestier jusqu'à un continent. Dès lors que cette fragmentation intervient dans des milieux très contrastés, l'adaptation des deux populations va être activée tandis que le transfert d'information sera inhibé. La spéciation est alors stimulée. Ainsi le monde végétal se caractérise par une forte capacité de colonisation, une forte capacité de transfert d'information génétique, une forte plasticitéplasticité morphologique, écologique, biologique, chronologique... Toutes ces propriétés s'appuient sur le principe de la réponse au changement ce qui fait que le monde végétal est parfaitement adapté à l'évolution des conditions du milieu.
2 - Conclusions
Ainsi contrairement aux apparences, la plante est un être vivant très mobilemobile. En plus des déplacements des individus ou du patrimoine génétique, on peut évoquer les mouvements physiologiques qui s'effectuent en permanence à l'intérieur d'une plante (nasties), mais également le mouvement des vrilles chez certaines lianes qui leur permettent de trouver un point d'accroche pour leur développement volubile. Notons également le mouvement des feuilles chez certaines légumineuseslégumineuses qui s'ouvrent le matin et se replient le soir (photonastie), ou chez les plantes dîtes sensitives qui sont capables de refermer leur feuillage instantanément sous la pressionpression du toucher (seismonastie). Les plantes sont bien mobiles. Cependant, les mouvements des végétaux sont soit trop lents, soit trop limités dans l'espace pour être directement perceptibles par l'homme; tout se passe comme s'ils se déroulaient dans une espace-tempsespace-temps auquel l'homme et les animaux en général, n'ont pas accès.
Quel peut être le rapport intérêt/risque dans la dispersion. Faut il mieux occuper un espace que l'on connaît ou risquer sa progéniture vers des lieux inconnus ?
Ces déplacements offrent aux végétaux les moyens de coloniser de nouveaux espaces, de sortir de l'influence de la plante mère. Bien entendu certains déplacements sont létaux (une spore de fougèrefougère qui tombe dans le désertdésert ! une graine de jacinthe d'eau qui tombe en haut d'une montagne,....), mais d'autres sont au contraire favorables. Seul le nombre des mouvements permet statistiquement d'atteindre tous les milieux différents. Les plantes ne mettent pas toutes leur destinée dans l'une ou l'autre des stratégies et bien généralement elles adoptent la prudence, c'est à dire se disperser loin et assurer parallèlement la colonisation à petites distances. D'ailleurs si elles ne savaient que se disperser loin elles diminueraient les probabilités statistiques de se rencontrer : un arbrearbre pousserait et ne produirait sa descendance qu'éloigné de lui....
Ces caractéristiques confèrent aux espèces végétales une grande capacité évolutive qui se traduit notamment par une adaptation perpétuelle de la végétation aux conditions locales du milieu.















