

au sommaire
Durant la méiose se produisent des échanges entre les chromosomes d'une même paire, les crossing-over, avant que ces chromosomes ne se séparent. En étudiant chez la plante modèle Arabidopsis thalianaArabidopsis thaliana un gène déjà connu chez la levurelevure, les chercheurs de l'INRA de Versailles, ont montré qu'il existait chez les plantes comme chez la levure deux types de crossing-over distincts. De plus, cette étude confirme qu'une des fonctions majeures des crossing-over est de permettre la bonne séparationséparation des chromosomes d'une même paire durant la méiose. Les données actuelles suggèrent que le brassage génétique qui se produit durant la méiose ne serait à l'origine qu'un effet collatéralcollatéral, qui se serait conservé au cours de l'évolution grâce à l'avantage sélectif qu'il procure. Ces résultats sont publiés le 26 Avril 2005 dans la revue "Current Biology".


© MAITRE Christophe - INRA
Plante expérimentale : Arabidopsis Thaliana.
Tous droits réservés
Le gène MER3 est connu chez la levure pour son rôle dans la formation des crossing-over au cours de la méiose. Le même gène existant chez les plantes, les chercheurs ont étudié un mutant d' Arabidopsis thaliana chez lequel ce gène ne fonctionne pas. Ces mutants donnent trois fois moins de graines. Les chercheurs ont observé que chez ces mutants, durant la méiose, près de 80% des crossing-over ne se produisent pas. Pour certaines paires, aucun crossing-over n'est observé, ce qui n'arrive jamais chez les plantes non mutées. En l'absence de crossing-over entre les deux chromosomes d'une paire, les chercheurs ont vérifié que, comme chez la levure, la répartition des chromosomes d'une même paire était aléatoire (cf. photo J), ce qui affecte fortement la production des gamètesgamètes (ovulesovules et grains de pollenpollen). Une fonction majeure du crossing-over est donc bien de permettre la méiose.
 Raphaël Mercier, INRA")
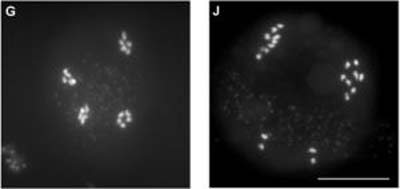
Photo G : répartition des chromosomes entre les quatre cellules issues d'une méiose normale chez Arabidopsis thaliana : chaque cellule sexuelle a bien reçu cinq chromosomes. (C) Raphaël Mercier, INRA
La recombinaisonrecombinaison (1) chez les mutants MER3 n'est cependant pas nulle : en utilisant des marqueurs génétiques, les chercheurs ont confirmé qu'il existe, comme chez la levure, une autre voie biochimique d'apparition des crossing-over, contrôlée par d'autres gènes. Les chercheurs ont de plus montré que ces deux voies sont très notablement différentes. Les crossing-over contrôlés par le gène MER3 ne peuvent être très proches l'un de l'autre sur le chromosome. Ce phénomène, dit d'interférenceinterférence, engendre une distribution régulière des crossing-over sur l'ensemble des chromosomes et assure que chaque paire de chromosomes subisse au moins un crossing-over. Ceci garantit la bonne séparation des chromosomes lors de la méiose. A l'inverse, dans la deuxième voie, les crossing-over se répartissent au hasard.


© WEBER Jean - INRA
Serre de collection des écotypesécotypes d'Arabidopsis thaliana à l'Unité de Recherche de Versailles.
Tous droits réservés
L'existence de deux voies de production des crossing-over se retrouve chez la levure, les plantes et probablement chez les mammifèresmammifères (où les deux types de gènes impliqués existent). Le ver nématodenématode C. elegans ne possède que la voie contrôlée par MER3. La levure S. pombe est différente : elle ne possède que la deuxième voie, celle où les crossing-over se répartissent au hasard. N'ayant que 3 chromosomes, la probabilité de ne pas avoir de crossing-over sur l'un d'eux reste cependant faible.
D'un point de vue évolutif, il est risqué pour un être vivant de ne posséder qu'une seule copie de son information génétique. L'apparition de cellules à 2N chromosomes a donc été un avantage sélectif au cours de l'évolution. La division par deux du nombre de chromosomes dans les cellules sexuelles permet d'éviter son augmentation sans fin. Grâce au mécanisme de crossing-over interférant auquel MER3 participe, au moins un crossing-over se produit sur chaque chromosome, ce qui assure une méiose régulière. Le brassage génétique résultant apparaît ainsi comme fortuit et secondaire, au moins initialement. L'avantage sélectif qu'il a très probablement conféré aux organismes sexuéssexués explique qu'il ait été conservé au cours de l'évolution.
(1) Chez la plupart des êtres vivants, le cycle de vie présente deux phases : une où les cellules de l'organisme portent un double jeu de chromosomes (noté 2N) et l'autre où elles n'en n'ont qu'un seul (noté N). Lors de la fécondationfécondation, deux cellules à N chromosomes fusionnent pour donner une cellule à 2N chromosomes. La méiose est le phénomène inverse, où une cellule à 2N chromosomes va se diviser deux fois pour donner quatre cellules à N chromosomes. Lors de la première division, les chromosomes homologueschromosomes homologues de chacun des deux jeux s'apparient par paire. Ils échangent des portions équivalentes de chromosomes au cours de crossing-over. Cette recombinaison assure un brassage génétique, puis les paires se dissocient pour donner deux cellules à N chromosomes. Il se produit toujours au moins un crossing-over par chromosome. La deuxième division est une division cellulaire classique. (cf. photo G)

par INRA
le 7 mai 2005



 un parasite des arbres. © David R. Tribble - CC BY-NC 3.0")











. L’iris possède une ouverture en son centre : la pupille, ronde, qui laisse passer la lumière. Le diamètre de la pupille peut varier, laissant plus ou moins entrer la lumière, en fonction de la luminosité ambiante. L’iris contient un pigment brun plus ou moins abondant.
© Mattis2412, CC by-sa 3.0")







